Česká a slovenská psychiatrie

Časopis
Psychiatrické společnosti ČLS JEP
a Psychiatrickej spoločnosti SLS
souborný článek / review article
- AKTUÁLNÍ ČÍSLO
- ARCHIV
- VYHLEDÁVÁNÍ
- PERIODIKUM
- REDAKČNÍ RADA
- PŘEDPLATNÉ
- INZERCE
- AKTUALIZOVANÉ
POKYNY
PRO AUTORY
- ISSN 1212-0383




- © Česká a Slovenská psychiatrie 2024
- © Galén 2024
VYBRANÉ GENETICKÉ PARAMETRY V ETIOPATOGENEZI DEPRESIVNÍ PORUCHY A ALZHEIMEROVY CHOROBY
SELECTED GENETIC PARAMETERS IN THE ETIOPATHOGENESIS OF DEPRESSIVE DISORDER AND ALZHEIMER
Eva Kitzlerová1, Zdeněk Fišar1, Jiří Raboch1
1 Psychiatrická klinika 1. LF UK a VFN, Praha
Práce byla podpořena grantem PRVOUK- P26/LF1/4 a projektem MZ-ČR-RVOVFN 64165.
SOUHRN
Kitzlerová E, Fišar Z, Raboch J. Vybrané genetické parametry v etiopatoge-nezi depresivní poruchy a Alzheimerovy choroby
Při poruchách nálady a neurodegenerativních chorobách nebyly identifikovány geny s přímým patogenetickým vlivem, ale existuje významný genetický podíl pro vznik jak depresivní poruchy, tak Alzheimerovy choroby. Deprese je rizikový faktor, prodrom a doprovodný symptom Alzheimerovy choroby. Mezi geny spojované s rizikem vzniku depresivní poruchy patří SLC64A pro serotoninový transportér (5-HTT), HTR2A pro serotoninový receptor 2A (5-HT2A), TPH2 pro neuronální tryptofanhydroxylázu 2, MAOA pro monoaminooxidázu A, COMT pro katechol-O-metyltransferázu, BDNF pro mozkový neurotrofní faktor, APOE pro apolipoprotein E (ApoE) a další. Hlavním genetickým rizikovým faktorem Alzheimerovy choroby s pozdním nástupem je výskyt ?4-alely APOE. Předložený text přináší přehled faktů o vybraných zkoumaných genetických parametrech a jejich úloze v etiopatogenezi poruch nálady a neurodegenerativních chorob.
Klíčová slova: genetika, depresivní porucha, Alzheimerova choroba
SUMMARY
Kitzlerová E, Fišar Z, Raboch J. Selected genetic parameters in the etiopathogenesis of depressive disorder and Alzheimer's disease
Genes have not been identified in mood disorders and neurodegenerative diseases with a direct pathogenetic effect, but the impact of genetic factors is very important in both disorders. Depression is a risk factor, prodromal and also the accompanying symptom of Alzheimer's disease. Genes associated with depression are SLC64A for serotonin transporter (5-HTT), HTR2A for serotonin receptor 2A (5-HT2A), TPH2 for neuronal tryptophanhydroxylase 2, MAOA for monoamine oxidase A, COMT for catechol-O-metyltransferase, BDNF for brain-derived neurotrophic factor, APOE for apolipoprotein E (ApoE) etc. The main genetic risk factor for the development of Alzheimer's disease with the late onset is ?4-alelle of APOE. This review brings the summary of facts about selected examined genetic parameters and their roles in the etiopathogenesis of affective disorders and neurodegenerative disorders.
Key words: genetics, depressive disorder, Alzheimer
ÚVOD
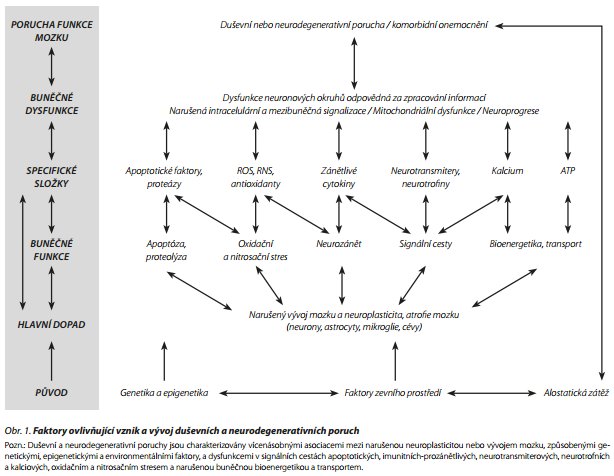
Etiologie duševních poruch je komplexní. Obecně se na ní podílejí faktory genetické a vlivy zevního prostředí. Rozlišuje se korelace genů a prostředí, kdy genetické vlivy určují, zda je subjekt danému prostředí vystaven, a interakce genů a prostředí, kdy geny určují náchylnost jedince k vlivům zevního prostředí1 a zevní vlivy způsobují epigenetické změny. Na etiopatogenezi některých duševních chorob, jako např. u deprese, demence či schizofrenie, se podílejí faktory genetické, epigenetické (metylace DNA, acetylace histonů), vlivy prostředí (např. těžké kovy, toxiny, kouření cigaret, dieta, cvičení aj.) a další proměnné (demografické - věk, pohlaví, porodní komplikace, zneužívání v dětství, úrazy hlavy či stresující životní události).2 Jen vzácně je příčina duševní nemoci jednoduchá (obr. 1). Při poruchách nálady a neurodegenerativních chorobách nebyly identifikovány geny s přímým patogenetickým vlivem. Deprese je rizikový faktor, prodrom a doprovodný symptom Alzheimerovy choroby.3 O míře heretability (vlivu genetických faktorů na přítomnost či nepřítomnost duševního onemocnění) informuje tzv. Holzingerův index. Pokud by duševní porucha byla podmíněna výhradně geneticky a faktory prostředí by neměly žádný vliv, heretabilita by byla rovna jedné. Poruchy nálady mají např. heretabilitu v rozmezí 0,4-0,6, demence 0,4-0,7, schizofrenie 0,6-0,8.4
Model depresivní poruchy zahrnuje faktory genetické a faktory vnějšího prostředí v průběhu života a naznačuje tak heterogenní etiologii depresivní poruchy. Vlivy vnějšího prostředí v raném věku ovlivňují adaptibilitu genomu vůči faktorům vnějšího prostředí v pozdějším věku. Zatímco samotná diagnóza depresivní poruchy je slabým prediktorem responze na specifickou léčbu, podtypy podle etiologie mohou pomoci ve výběru mezi léčebnými možnostmi. Tato hypotéza byla navržena s ohledem na responzi jednotlivých, etiologických podtypů deprese k psychologické léčbě či antidepresivní léčbě.5 Rané zkušenosti s rodičovskou péčí nebo jejím odmítnutím mají vliv na rozvoj depresivní poruchy v dospělosti.6 Tyto sociální faktory ale nevysvětlují větší rozdíly v riziku vzniku deprese mezi monozygotními a dizygotními dvojčaty7, 8 Byla nalezena specifická interakce mezi funkčním polymorfismem v genu pro serotoninový transportér (5-HTTL-PR) a stresujícími životními událostmi. Abúzus v dětství a stresové životní události byly asociovány s vysokým rizikem rozvoje depresivní poruchy u osob s krátkými alelami, ale měly malý vliv na rozvoj deprese u homozygotů s dlouhými alelami.9 Obdobně i u dalších neurodegenerativních chorob nebyly dosud jasně a jednoznačně identifikovány specifické geny podílející se na rozvoji psychopatologie.10-12 Obecně, kandidátními geny jsou zejména geny pro neurotransmiterové receptory, jejichž polymorfismy následně ovlivňují funkci a aktivitu proteinů, které se různým způsobem podílejí na stavbě a funkci nervové soustavy.
S etiologií mnoha psychiatrických onemocnění jsou často mimo jiné spojovány poruchy serotonergní neurotransmise.13 Serotonin se v centrálním nervovém systému účastní především procesů, které se podílejí na vzniku, rozvoji a modulaci nálad. Jeho nedostatek je spojován nejčastěji s depresí a může vést také až k podrážděnosti a zvýšené agresivitě.14 Změny v metabolismu serotoninu jsou pravděpodobně odpovědné za řadu psychických poruch nebo jejich symptomů. Potenciace serotonergního systému v mozku ovlivňuje též významně kognitivní procesy, resp. kognitivní flexibilitu.14, 15 Serotonin má významný vliv i mimo centrální nervový systém v celé řadě dalších fyziologických procesů. Ovlivňuje např. svalový tonus, podporuje kontrakce hladkého svalstva a krevní srážlivost, proto řada psychických poruch souvisejících s poruchami serotoninového transportu může být přímo spojena s interními a kardiologickými poruchami.14
U Alzheimerovy choroby nejsou vlastní příčiny této závažné choroby stále zcela jasné kromě 1-5 % případů s rozpoznanými genetickými rozdíly. Jisté je, že jde o chorobu multifaktoriální, na jejímž vzniku se podílí řada faktorů (obecně faktorů dědičných i nedědičných). Choroba postihuje muže i ženy stejně často, celkově může být více pacientů ženského pohlaví, neboť jedním z rizikových faktorů Alzheimerovy choroby je i věk a právě ženy se průměrně dožívají vyššího věku než muži. Z nedědičných faktorů se dále uvádí například obezita, hypertenze, některé infekce a cévní faktory. Z genetických faktorů se nejčastěji uvádí mutace ve čtyřech hlavních genech: APP (kódující amyloidový prekurzorový protein; 21q21.2-21q21.3), APOE (kódující apolipoprotein E; 19ql3.2), PSEN1 (kódující presenilin 1; 14q24.3) a PSEN2 (kódující presenilin 2; Iq31-q42).15 Uvádí se celá řada dalších genů, které by mohly mít s Alzheimerovou chorobou větší či menší souvislosti (obecně se označují AD5-AD16).
Podle doby vzniku a rychlosti nástupu se rozlišuje varianta Alzheimerovy nemoci familiární a sporadická. Familiární forma s časným nástupem ("early onset familial") začíná před 65 lety věku, je vzácná a je obvykle spojena s mutacemi v genech APP, PSEN1 nebo PSEN2. Sporadická varianta s pozdním nástupem ("late onset sporadic") má jako rizikové faktory věk, vlivy zevního prostředí a výskyt ?4 alely genu APOE.
Na Psychiatrické klinice 1. LF UK a VFN v Praze byly v rámci projektu PRVOUK a Výzkumného záměru "Neuropsychiatrické aspekty neurodegenerativních chorob" u pacientů trpících depresivní poruchou, Alzheimerovou chorobou a u zdravých kontrol měřeny parametry biochemické, klinické a zároveň byly provedeny analýzy některých genetických parametrů. Obecným cílem výzkumu bylo nalezení biologických markerů, které by mohly hrát důležitou roli v etiopatogenezi neurodegenerativních chorob. Cílem dílčího hodnocení genetických parametrů byla analýza asociace polymorfismů vybraných genů s výskytem depresivní poruchy a Alzheimerovy demence a možnosti využití potenciálních interakcí (kombinovaného vlivu) těchto polymorfismů ve vztahu k riziku vzniku depresivní poruchy a Alzheimerovy demence. Testována byla dále hypotéza, že depresivní porucha a Alzheimerova demence mohou mít částečně společný genetický základ. Analyzovány byly polymorfismy genů pro serotoninový transportér (5-HTT), mozkový neurotrofní faktor (BDNF), proteiny teplotního šoku (HSP70), serotoninový receptor 2A (5-HT2A) a apolipoprotein E (ApoE). O hodnocených genetických parametrech pojednává přehled níže.
SEROTONINOVÝ TRANSPORTÉR (5-HTT)
Nejdůležitější gen kontrolující funkci serotoninu (5-hydroxytryptamin, 5-HT) je gen pro 5-HTT (SLC6A4). Lidský gen SLC6A4 (chromozomální umístění 17q11.2) kóduje integrální membránový protein, který transportuje serotonin přes membránu a zajišťuje zpětné vychytávání serotoninu v chemických synapsí.9 Spolu s geny pro monoaminooxidázu typu A (MAOA) a katechol-O-metyltransferázu (COMT) je SLC6A4 je jedním z nejvíce studovaných genů ve vztahu k narušené monoaminové neurotransmisi při poruchách nálady a neurodegenerativních onemocněních. Lidské genetické varianty spojené s genem SLC6A4 zahrnují dva nejčastěji studované polymorfismy:
1. Délkové varianty v polymorfní oblasti spojené s genem pro serotoninový transportér (5-HTTLPR, "serotonin transporter genelinked polymorphic region") ; krátká varianta (S) má 14 repeticí sekvence, zatímco dlouhá varianta (L) má 16 repeticí.16 Krátká varianta vede ke snížení transkripce genu SLC6A4. Předpokládá se, že polymorfismus nacházející se v 5-HTTLPR má vliv nachování.17
2. Tandemové repetice s proměnným počtem opakování (VNTR, "variable number of tandem repeats") ve funkčním druhém intronu (STin2 VNTR); polymorfismus byl zjištěn se třemi alelami: STin2.9, STin2.10 a STin2.12 (9, 10 a 12 opakování).VNTR mohou podle svého počtu ovlivňovat expresi genů. VNTR polymorfismus byl spojen s afektivními poruchami18 a schizofrenií.19
Nejprve byl studován vztah mezi polymorfismem 5-HTTLPR lidského SLC6A4 a rysy spojenými s úzkostnými stavy20 Homozygoti se dvěma alelami S nebo heterozygoti jsou ve srovnání s homozygoty pro alelu L více senzitivní k úzkostným podnětům.21, 22 Byl učiněn závěr, že 5-HTTLPR může mít malý, ale hodnověrný vliv na osobnost, zejména v projevu rysů úzkosti.23 Průlomovou prací týkající se vlivu polymorfismů genu serotoninového transportéru na rozvoj deprese a sebevražedného jednání po nepříznivých životních událostech byla práce Caspiho et al. z roku 2003,9 podle níž jedinci s jednou nebo dvěma kopiemi krátké alely promotorového polymorfismů SLC6A4 vykazovali více příznaků deprese a sebevražedné sklony vzhledem k stresující životní události než jednotlivci homozygotní pro dlouhou alelu. Genetické varianty 5-HTTLPR byly studovány také jako prediktor antidepresivní odpovědi. Byl učiněn závěr, že u bělochů může být 5-HTTLPR prediktorem antidepresivní odpovědi a remise.24
I když aktivita 5-HTT v mozku by mohla být spojena s polymorfismy 5-HTTLPR, není stále jasné, zda aktivita 5-HTT reguluje spojení mezi stresem a depresivní poruchou. Některé metaanalýzy podporují hypotézu, že polymorfismus 5-HTTLPR reguluje vztah mezi stresem a depresí,25 jiné tuto hypotézu nepodporují.26, 27 Několik studií našlo spojení mezi antisociálním a agresivním chováním a výskytem varianty S polymorfismu 5-HTTLPR; nicméně tato asociace nebyla přesvědčivě prokázána.28
Řadu konfliktních a nekonzistentních výsledků v této oblasti lze vysvětlit zanedbáním vlivu etnika a vlivu dalších jednonukleotidových polymorfismu (SNP), které mění funkční účinky L a S varianty.29 Existuje totiž více funkčních variant lidského SLC6A4,16, 30 které vznikají v důsledku alternativních promotorů, diferenciálního sestřihu (zahrnujícího exony 1A, IB a 1C), variability 3'-nepřekládané oblasti (UTR, "3'-untranslated-region") a dalších SNP.31 Alely (S, L) a genotypy (LL, LS, SS) 5-HTTLPR by měly být zkoumány v kombinaci s polymorfismem VNTR a se dvěma úzce spojenými SNP (rs25531, rs25532), které mění expresi, transport a funkci 5-HTT. Například polymorfismus rs25531 spočívá v záměně nukleotidů A > G, která dává LA a LG alely. Frekvence alel jsou u různých etnik velmi rozdílné; v bělošské populaci má polymorfismus rs25531 frekvenci alely LA asi 50 % a alely LG asi 13 %. Protože je LG funkčně ekvivalentní alele S,32 významná část nositelů L alely může být funkčně ekvivalentní s nositeli S alely, která je spojena s nižší úrovní mRNA serotoninového transportéru a nižší expresí 5-HTT v lidských buněčných liniích.29
Ačkoliv byl polymorfismus 5-HTTLPR poměrně podrobně zkoumán ve vztahu k faktorům vnějšího prostředí, zdá se nepravděpodobné, aby tento gen působil izolovaně. Polymorfismus na minimálně dvou dalších kandidátních genech vykazoval podobné interakce s faktory vnějšího prostředí. SNP v genu kódujícím BDNF (BDNF) interagoval s 5-HTTLPR na molekulární a systémové úrovni 33 a dále posiloval vztah mezi 5-HTTLPR a environmentální adverzitou v epidemiologických studiích.34-36 Lidské epidemiologické studie naznačují, že BDNF 66 met alela interaguje s 5-HTTLPR krátkou alelou a určuje citlivost vůči pozitivním a negativním vnějším faktorům. Tato trojitá interakce může být užitečná při léčbě deprese, zvláště pak, když BDNF ovlivňuje efekt antidepresiv, především těch, které inhibují serotoninový transportér.36-38 Pokud se vezme ještě v úvahu dysregulace hypotalamo-hypofyzo-adrenální osy (HPA), jednak ve spojitosti s nepříjemnými zkušenostmi životními a jednak s depresí,39, 40 pak není žádným překvapením, že gen kódující hormon uvolňující kortikotropin (CRHR1) se vyskytuje též v souvislosti s dětským abúzem při vzniku deprese v dospělosti.41 SLC6A4, BDNF ani CRHR1 geny nejsou přímo asociovány s depresivní poruchou. Polymorfismus v některých genech se může kombinovat a vést k dispoziční citlivosti či rezistenci vůči faktorům vnějšího prostředí bez přímého vlivu na zvýšení rizika vzniku deprese.
Polymorfismus 5-HTTLPR má vliv na expresi serotoni-nového transportéru a tím i na účinek antidepresiv a může tak ovlivnit odpovědna léčbu. Depresivní nemocní s variantou L vykazují lepší odpovědna léčbu SSRI,42,43 zatímco krátká varianta S je spojena s pomalejší odpovědí na běžnou antidepresivní léčbu.44, 45 Jedinci s alelou S mohou mít zvýšené riziko výskytu nežádoucích reakcí na léčbu (jako jsou závratě, poruchy spánku, únava, GIT obtíže).
Polymorfismus genu SLC6A4 je nejčastěji sledován v souvislosti s odpovědí na léčbu antidepresivy a jsou velmi málo známy souvislosti s jinými léčivy. 13 Přesto existují data uvádějící jeho souvislost i s atypickými antipsychotiky.46 Ve studii s 227 ambulantními pacienty a 420 zdravými kontrolními subjekty bylo zjištěno, že kombinace polymorfismu rs6311 genu pro 5-HT2A receptor a variantou S polymorfní alely 5-HTTLPR genu pro 5-HTT byla asociována se schizofrenií.47 Polymorfismus 5-HTTLPR ovlivňuje i odpovědna léčbu risperidonem, jak prokázala studie na vzorku 129 čínských pacientů, L-alela 5-HTTLPR se vyskytuje signifikantně méně u non-respondérů v porovnání s respondéry. Odpovědna léčbu byla hodnocena škálou BPRS (Brief Psychiatrie Rating Scale), která zahrnuje nejdůležitější položky škály PANSS. Analýza polymorfismu LPR a VNTR ukázala, že haplotyp L-12 je spojen s lepší odpovědí na léčbu risperidonem.48 Týž závěr podporuje i brazilská studie s klozapinem na 116 pacientech evropského původu: S-alela polymorfismu 5-HTTLPR je spojena s horší odpovědí na léčbu (opět hodnoceno škálou BPRS) v porovnání s L-alelou.49
SEROTONINOVÝ RECEPTOR 2A (5-HT2A)
Polymorfismus genu 5HT2A pro serotoninový receptor 5-HT2A se nachází v promotorové oblasti genu 5HT2A. Polymorfismus 5HT2A byl studován ve vztahu k léčebné odpovědi u schizofrenie a jeví se nadějně ve vztahu k predikci terapeutické odpovědi na antipsychotika druhé generace. Vesměs se výsledky větších studií shodují v tom, že polymorfismy 5HT2A mohou přispívat k variabilitě odpovědi na léčbu klozapinem, ale pouze v interakci více polymorfismu genů se vztahem k neurotransmiterům.47, 50 Méně často byla studována souvislost mezi odpovědí na léčbu a polymorfismy jiných podtypů serotoninových receptorů(5-HT2C, 5-HT3).51 V animálních studiích bylo prokázáno snížení exprese genů 5HT2A a 5HT2C po podávání klozapinu, haloperidolu a olanzapinu.52, 53
Se serotoninovými 5-HT2A receptory je spojován též výskyt vedlejších účinků antidepresiv na sexuální funkce. A/A genotyp 5HT2A může být genetickým znakem pro časnou detekci sexuálních dysfunkcí vyvolaných serotonergními antidepresivy u mladých mužů s těžkou depresivní poruchou, zatímco G alela je protektivní.54
MOZKOVÝ NEUROTROFNÍ FAKTOR (BDNF)
BDNF je členem neurotrofní rodiny růstových faktorů. Neurotrofní faktory se nacházejí v mozku a na periferii.
BDNF působí na určité neurony v centrálním a periferním nervovém systému a pomáhá podporovat přežití stávajících neuronů, růst a diferenciaci nových neuronů a synapsí. V mozku je aktivní např. v hipokampu, mozkové kůře a předním mozku, oblastech, které jsou životně důležité pro učení, paměť a vyšší myšlení. Protein BDNF je kódován genem BDNF, který je u lidí lokalizován na chromosomu 11p13. Funkční in vitro studie BDNF ukázaly, že ovlivňuje intracelulární transport, sbalení a uvolnění proteinu. Myší model varianty genu BDNF vykazuje podobnost s lidským fenotypem a je asociovaný s úzkostným chováním a poruchou vyhasínání reakce na negativní zkušenost. Při depresi a zvýšeném stresu dochází k aktivaci stresové osy HPA, zvýšení stresových hormonů (např. kortizolu) a snížení BDNF, což se pravděpodobně podílí na úbytku synaptických spojů mezi neurony. Různé meta-analýzy potvrdily, že může být korelace mezi nízkou aktivitou BDNF a vznikem bipolární poruchy.55-57
Všechny farmakoterapeutické antidepresivní zásahy vedou ke zvýšení BDNF, dokonce i elektrokonvulze vedou ke zvýšení BDNF, stejně jako jeho receptoru trkB.58 Zdá se, že antidepresiva díky svým genomickým účinkům vykazují neuroprotektivní až neurotropní působení v hipokampu a v jiných strukturách mozku. Molekulární hypotéza deprese by pak mohla být podnětem pro vývoj nových postupů léčby deprese, především by mělo jít o látky stimulující signální systém cAMP - PKA (proteinkináza A) - CREB (protein vázající se v genomu na element zodpovědný za odezvu na cAMP, "cAMP response element binding protein") - BDNF.59
BDNF hraje klíčovou roli v patofyziologii poruchy nálady související se stresem. Akutní stres vyvolal rychlé zvýšení hladiny BDNF v séru, zatímco dlouhodobý stres vedl k depresi a ke snížení hladin BDNF.60 Úloha BDNF v patofyziologii duševních poruch byla doložena zvýšením sérových a mozkových koncentrací BDNF vlivem antidepresiv, stabilizátorů nálady a antipsychotik.61-63
Metaanalýza genetických asociačních studií při bipolární poruše ukázala, že významné mohou být polymorfismy genů pro BDNF, dopaminový receptor 4, aktivátor oxidáz D-aminokyselin a tryptofanhydroxylázy Nicméně žádný z těchto polymorfismů nebyl významný po korekci na vícenásobné testování.64 Val66Met polymorfismus genu BDNF je funkčně relevantní SNP, který ovlivňuje sekreci BDNF a podílí se na rozdílech v objemu hipokampu u neuropsychiatrických pacientů oproti kontrolám, i když metaanalýza neprokázala tuto asociaci mezi polymorfismem BDNF a objemy hipokampu.65 U pacientů s Alzheimerovou chorobou byly studovány polymorfismy Val66Met (G196A), C270T a 11757 G/C genu BDNF. Výsledky jsou rozporuplné.66 Polymorfismy 11757 G/C BDNF byly významně asociovány s Alzheimerovou chorobou.67 Frekvence alely 11757 C byla pozorována významně vyšší u pacientů s Alzheimerovou chorobou a depresí oproti pacientům s Alzheimerovou chorobou bez deprese. Nositelé A alely G196A odpovídali lépe na léčbu paroxetinem.68 U nosičů polymorfismů Val66Met jsou zjišťována větší depozita amyloidu beta (A?) v mozku, snížení objemu hipokampu, snížení výkonnosti epizodické paměti, tedy základní atributy Alzheimerovy choroby. Tento polymorfismus představuje nebezpečí zvláště v kombinaci s nositelstvím s4 alely APOE.69
PROTEINY TEPLOTNÍHO ŠOKU 70 (HSP70)
Proteiny teplotního šoku (HSP, "heat shock protein") je označení pro skupinu proteinů, které zajišťují univerzální stresovou odpověď buněk, například při vystavení buňky vysoké teplotě, extrémnímu pH nebo třeba těžkým kovům. Vážou se na nesbalené proteiny, chrání je či pomáhají jejich sbalení (chaperonová aktivita). Chaperon je speciální protein, který v buňce pomáhá skládat většinu bílkovin do jejich správného prostorového uspořádání. Tím tedy těmto proteinům umožňují vykonávat jejich funkci v buňce. Chaperony se vyskytují jak v eukaryotických buňkách, tak i u bakterií a v mitochondriích. HSP70 je jeden z nejběžnějších proteinů teplotního šoku pracující jako chaperon. HSP70 se po navázání ATP dostává do otevřené konformace, v níž je schopen vázat hydrofobní části špatně složených proteinů. Následně dochází k hydrolýze ATP a uzavření chaperonového komplexu, čímž protein dostane možnost sbalit se do správného prostorového uspořádání. V následující fázi dojde k výměně ADP za ATP a cyklus je možné opakovat. Pro správné složení jednoho proteinu je obvykle potřeba několikrát celý cyklus opakovat. HSP70 takto ovlivňuje konformaci širokého spektra polypeptidů a v buňce se objevuje ve zvýšeném množství tehdy, je-li buňka vystavená určitým formám stresu (např. teplotnímu šoku). Služeb HSP70 proteinů se ale využívá i při translokaci bílkovin do plastidů a mitochondrií, kde je zase nutné, aby přepravovaný protein měl spíše rozbalenou formu a neskládal se do své nativní konformace.
HSP70 proteiny chrání buňky před teplotním nebo oxidačním stresem, podílí se na likvidaci poškozených nebo vadných proteinů a HSP70 přímo inhibuje apoptózu, brání agregaci/oligomeraci A? a ? proteinu - působí neuroprotektivně, jsou schopny aktivovat přirozenou i specifickou imunitu, a to prostřednictvím regulace c-Jun N-terminalkinázy HSP72 ("heat shock 70kD protein 1A") je kódován genem HSPA1A. V rámci našeho výzkumu byly analyzovány 2 polymorfismy v genu HSPA1A, rsl043618(+190G/C)arsl008438(-110A/C).70
APOLIPOPROTEIN E (ApoE)
Apolipoprotein E (ApoE) je polymorfní glykoprotein s hmotností 34 kDa hrající důležitou roli v metabolismu plazmových lipidů a transportu cholesterolu. Podílí se na distribuci lipidů při vývoji a reparaci nervové tkáně. Funguje též jako imunomodulátor, regulátor růstu a diferenciace buněk. U člověka byly identifikovány tři jeho izoformy (ApoE2, ApoE3 a ApoE4), kódované třemi alelami (?2, ?3, ?4) genu APOE lokalizovaného na chromosomu 19. Tyto tři alely kódují rozdílnou informaci o lokalizaci aminokyseliny argininu a cysteinu v pozicích 112 a 158 peptidového řetězce ApoE.59 V posledních letech se dostalo velké pozornosti úloze APOE alel u neurodegenerativních chorob, např. přítomnost ?2 alely je spojována se snížením depresivních příznaků u pacientů s roztroušenou sklerózou, naopak ?4 alela vede ke zhoršení a časnějšímu nástupu depresí. Přítomnost ?4 alely je dále pokládána za rizikový faktor pro poškození kognitivních funkcí. Ve vztahu k Alzheimerově chorobě je zejména alela ?4, neboť nejvyšší riziko objevení se Alzheimerovy demence bylo pozorováno u genotypu 4/4 a nejnižší u genotypu 2/2, 2/3 a 3/3. Přítomnost alely ?4 posouvá začátek choroby asi o 5 let dopředu a patří mezi významné rizikové faktory Alzheimerovy choroby.71 Frekvence výskytu alely ?4 korelovala významně se skupinou demencí typu Alzheimerova demence nebo demencí s Lewyho tělísky nikoliv však s demencemi u Parkinsonovy choroby nebo Huntingtonovy chorey. Neuropatologická pozorování naznačují, že imunoreaktivita ApoE4 je asociována se senilními plaky dochází k interakci ApoE s APP a tato interakce je rozdílná u jednotlivých izoforem ApoE. Tomu odpovídá i skutečnost, že počet a denzita amyloidních plaků je zvýšená u pacientů s Alzheimerovou demencí s jednou nebo dvěma alelami ?4.72 ApoE4 reguluje kumulaci A(3, o biochemismu reakce nejsou informace známy. ApoE se též podílí na udržování integrity synapsí, a je tedy možné, že ApoE4 je z tohoto pohledu tou nejméně vhodnou izoformou, která vede k defektům na úrovni neuronů a jejich synapsí. ApoE4 též aktivuje reparační procesy na synapsích v procesu stárnutí či mechanického poškození a izoforma ApoE4 může být méně výkonná než ostatní dvě izoformy ApoE4 reguluje hladinu intracelulárního kalcia, které se podílí na regulaci aktivity kináz a fosfatáz, které jsou klíčové pro fosforylaci proteinů. Tau protein je u Alzheimerovy demence abnormálně fosforylován, což vede k destabilizaci mikrotubulů a poruchám axonálního transportu. ApoE může tedy přispívat k patogenezi Alzheimerovy choroby právě přes cytoskeletální interakce.73 Úloha ApoE v nervovém systému spočívá především v udržování synaptické integrity v procesu neurodegenerace, ovlivňuje stabilitu neuronálního cytoskeletu a je prekurzorem syntézy nových synaptických zakončení. Souvislost Alzheimerovy demence s genovým polymorfismem APOE je zřejmá a výskyt alely ?4 je významným rizikovým faktorem objevení se této choroby v časnějším věku.74 Vztah mezi genotypem APOE a dalšími biomarkery nebo neuropsychiatrickými symptomy je dosud nejasný.
ZÁVĚR
Výsledky hodnocení některých biochemických parametrů měřených v periferní krvi osob s depresivní poruchou a Alzheimerovou chorobou byly již publikovány7"7 Korelace genetických polymorfismů vybraných genů k výskytu a k riziku vzniku depresivní poruchy a Alzheimerovy choroby a jednotlivým diagnostickým subtypům a dále případná korelace s parametry biochemickými a klinickými budou publikovány v blízké době.
LITERATURA
- 1. Hosák L, Šilhán P, Hosáková J: Interakce genů a prostředí v Psychiatrii. Česká a slovenská psychiatrie 2013; 109 (6): 304-309.
- 2. Van Winkel R, Esquivel G, Kenis G et al. Genome-widefindings in schizophrenia and the role of gene-environmentinterplay CNS Neurosci Ther 2010; 16: el85-192.
- 3. Enache D, Winblad B, Aarsland D. Depression in dementia: epidemiology, mechanisms, and treatment. CurrOpin Psychiatry. 2011; 24 (6): 461-472.
- 4. Nurnberger JI, Berrettini W. Psychiatric Genetics. London: Chapman &HallMedical 1998: 164.
- 5. Uher R: The implicationsof gene-environment interactions in depression: will cause inform cure? Molecular Psychiatry 2008, 13: 1070-1078.
- 6. Brown GW, Craig TK, Harris TO. Parental maltreatment and proximal risk factors using the Childhood Experience of Care & Abuse (CECA) instrument: a life-course study of adult chronic depression-5. J AfFectDisord 2008; doi:10.1016/j.jad.2008.01.016.
- 7. McGuffin P, Katz R.Watkins S, Rutherford J. A hospital-based twin register of the heritability of DSM-IV unipolar depression. Arch Gen Psychiatry 1996; 53: 129-136.
- 8. Sullivan PF, Neale MC, Kendler KS. Genetic epidemiology of major depression: review and meta-analysis. Am J Psychiatry 2000; 157:1552-1562.
- 9. Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H et al. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 2003; 301: 386-389.
- 10. Arango V, Huang YY, Underwood MD, Mann JJ. Genetics of the serotonergic system in suicidal behavior. J Psychiatr Res 2003; 37 (5): 375-386.
- 11. Kendler KS, Eaves L (eds). Psychiatric Genetics (Review of Psychiatry Series, 24, 1; Oldham JM, Rib a MB, series editors). Washington, DC, American Psychiatric Publishing 2005.
- 12. Sysoeva OV, Maluchenko NV, Timofeeva MA, Portnova GV, Kulikova MV, Tonevitsky AG, Ivanitsky AM. Aggression and 5HTT polymorphism in females: Study of synchronized swimming and control groups. Int J Psychophysiol 2009; 72: 173-178.
- 13. Kuželová H, Ptáček R, Macek M: Polymorfismus genu pro serotoninový transportér 5-HTT v etiologii psychiatrických onemocnění. Česká a slovenská psychiatrie 2010; 106 (2): 98-104.
- 14. Berger M, Gray JA, Roth BL. The expanded biology of serotonin. Ann Rev Med 2009; 60(1): 355-366.
- 15. Ridge PG, MTW, Kauwe JSK: Genetics of Alzheimer's Disease. Bio Med Research International 201; (Volume 2013): 1-13.
- 16. Nakamura M, Ueno S, Sano A, Tanabe H. The human serotonin transporter gene linked polymorphism (5-HTTL-PR) shows ten novel allelic variants. Mol Psychiatry. 2000; 5 (1): 32-8.
- 17. Heils A, Teufel A, Petri S, Stöber G, Riederer P, Bengel D, Lesch KR Allelic variation of human serotonin transporter gene expression. J Neurochem 1996; 66 (6): 2621-2624.
- 18. Anguelova M, Benkelfat C, Turecki G. A systematic review of association studies investigating genes coding for serotonin receptors and the serotonin transporter: I. Affective disorders. Mol Psychiatry 2003; 8 (6): 574-591.
- 19. Fan JB, Sklar P. Meta-analysis reveals association between serotonin transporter gene STin2 VNTR polymorphism and schizophrenia. Mol Psychiatry. 2005; 10 (10): 928-938, 891.
- 20. Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S, Benjamin J, Muller CR, Hamer DH, Murphy DL. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 1996; 274 (5292): 1527-1531.
- 21. Hariri AR et al. Serotonin transporter genetic variation and the response of the human amygdale. Science 2002; 297: 400-403.
- 22. http://gendep.iop.kcl.ac.uk/index.php - GENDEP study.
- 23. Schinka JA, Busch RM, Robichaux-Keene N. A meta-analysis of the association between the serotonin transporter gene polymorphism (5-HTTLPR) and trait anxiety. Mol Psychiatry 2004; 9 (2): 197-202.
- 24. Porcelli S, Fabbri C, Serretti A. Meta-analysis of serotonin transporter gene promoter polymorphism (5-HTTLPR) association with antidepressant efficacy Eur Neuropsychopharmacol 2012; 22 (4): 239-258.
- 25. Karg K, Burmeister M, Shedden K, Sen S. The serotonin transporter promoter variant (5-HTTLPR), stress, and depression meta-analysis revisited: evidence of genetic moderation. Arch Gen Psychiatry 2011; 68 (5): 444-454.
- 26. Munafó MR, Durrant C, Lewis G, Flint J. Gene' environment interactions at the serotonin transporter locus. Biol Psychiatry 2009; 65 (3): 211-219.
- 27. Risch N, Herrell R, Lehner T, Liang KY, Eaves L, Hoh J, Griem A, Kovacs M, Ott J, Merikangas KR. Interaction between the serotonin transporter gene (5-HTTLPR), stressful life events, and risk of depression: a meta-analysis. JAMA 2009; 301 (23): 2462-2471.
- 28. Tielbeek JJ, Karlsson Linnér R, Beers K, Posthuma D, Popma A, Polderman TJ. Meta-analysis of the serotonin transporter promoter variant (5-HTTLPR) in relation to adverse environment and antisocial behavior. Am J Med Genet B Neuropsychiatr Genet 2016; Mar 16. doi: 10.1002/ajmg.b.32442.
- 29. Murphy DL, Maile MS, Vogt NM. 5HTTLPR: White Knight or Dark Blight? ACS Chem Neurosci 2013; 4 (1): 13-15.
- 30. Murphy DL, Lesch KR Targeting the murine serotonin transporter: insights into human neurobiology Nat Rev Neurosci 2008; 9 (2): 85-96.
- 31. Murphy DL, Moya PR. Human serotonin transporter gene (SLC6A4) variants: their contributions to understanding pharmacogenomic and other functional GxG and GxE differences in health and disease. Curr Opin Pharmacol 2011; 11 (1): 3-10.
- 32. Hu XZ, Lipsky RH, Zhu G, Akhtar LA, Taubman J, Greenberg BD, Xu K, Arnold PD, Richter MA, Kennedy JL, Murphy DL, Goldman D. Serotonin transporter promoter gain-of-function genotypes are linked to obsessive --compulsive disorder. Am J Hum Genet 2006; 78 (5): 815-826.
- 33. Martinowich K, Lu B. Interactionbe-tween BDNF and serotonin: role in mooddisorders. Neuropsychopharmacology 2008; 33: 73-83.
- 34. Kaufman J, Yang BZ, Douglas-Palumberi H, Grasso D, Lipschitz D, Houshyar S et al. Brain-derived neurotrophic factor-5-HTTLPR gene interactions and environmental modifiers of depression in children. Biol Psychiatry 2006; 59: 673-680.
- 35. Kim JM, Stewart R, Kim SW, Yang SJ, Shin IS, Kim YH et al. Interactions between life stressors and susceptibility genes. Implications of G_E in depression (5-HTTLPR and BDNF) on depression in Koreanelders. Biol Psychiatry 2007; 62: 423-428.
- 36. Wichers M, Kenis G, Jacobs N, Mengelers R, Derom C, Vlietinck R et al. The BDNF Val66Met x 5-HTTLPR x child adversity interaction and depressive symptoms: An attempt at replication. Am J Med Genet B: Neuropsychiatr Genet 2008; 147: 120-123.
- 37. Di Bella D, Erzegovesi S, Cavallini MC, Bellodi L. Obsessive-CompulsiveDisorder, 5-HTTLPR polymorphism and treatment response. Pharmacogenomics J 2002; 2: 176-181.
- 38. Faraone SV, Khan SA. Candidate gene studies of attention-deficit/hyperactivity disorder. J Clin Psychiatry 2006; 67 (8): 13-20.
- 39. Fink LA, Bernstein D, Handelsman L, Foote J, Lovejoy M. Initial reliability and validity of the childhood trauma interview: a new multidimensional measure of childhood interpersonal trauma. Am J Psych 1995; 152 (9): 1329-1335.
- 40. Fišar Z et al. Vybrané kapitoly z biologické psychiatrie. Praha: Grada 2009.
- 41. Fumeron F, Betoulle D, Aubert R, Herbeth B, Siest G, Rigaud D. Association of a functional 5-ht transporter gene polymorphism with anorexia nervosa and food intake. Mol Psychiatry 2001; 6: 9-10.
- 42. Murphy DL, Lerner A, Rudnick G, Lesch KR Serotonin transporter: Gene, genetic disorders, and pharmacogenetics. Mol Interv 2004; 4: 109-123.
- 43. Pollock BG, Ferrell RE, Mulsant BH, Mazumdar S, Miller M, Sweet RA. Allelic variation in the serotonin transporter promoter affects onset of paroxetine treatment response in late-life depression. Neuropsychopharmacology 2000; 23: 587-590.
- 44. Benedetti F, Serretti A, Colombo C, Campori E, Barbini B, di Bella D. Influence of a functionalpolymorphism-withinthepromoterofthe serotonin transporter gene on the effects of total sleep deprivation in bipolar depression. Am J Psychiatry 1999; 156: 1450-1452.
- 45. Racine SE, Culbert KM, Larson CL, Klump KL. The possible influence of impulsivity and dietary restraint on associations between serotonin genes and bingeeating. J Psychiatr Res 2009; 43 (16): 1278-1286.
- 46. Bolonna AA, Arranz MJ, Mancama D, Kerwin RW. Pharmacogenomics - can genetics help in the care of psychiatric patients? Int Rev Psychiatry 2004; 16 (4): 311-319.
- 47. Saiz PA, Garcia-Portilla MP, Arango C et al. Association study of serotonin 2A receptor (5-HT2A) and serotonin transporter (5-HTT) gene polymorphisms with schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry 2007; 31 (3): 741-745.
- 48. Wang L, Yu L, He G et al. Response of risperidone treatment maybe associated with polymorphisms of HTT gene in Chinese schizophrenia patients. Neurosci Lett 2007; 414 (1): 1-4.
- 49. Kohlrausch FB, Salatino-Oliveira A, Gama CS et al. Influence of serotonin transporter gene polymorphisms on clozapine response in Brazilian schizophrenics. J Psychiatr Res 2010; 44 (16): 1158-1162.
- 50. Arranz MJ, Munro J, Birkett J et al. Pharmacogenetic prediction of clozapine response. Lancet 2000; 355 (9215): 1615-1616.
- 51. Souza RP, de Luca V, Meltzer HY, Lieberman JA, Kennedy JL. Influence of serotonin 3A and 3B receptor genes on clozapine treatment response in schizophrenia. Pharmacogenet Genomics 2010; 20 (4): 274-276.
- 52. Huang XF, Tan YY, Huang X, Wang Q. Effect of chronic treatment with clozapine and haloperidol on 5-HT2A and 2C receptor mRNA expression in the rat brain. Neurosci Res 2007; 59 (3): 314-321.
- 53. Padin JF, Rodriguez MA, Dominguez E et al. Parallel regulation by olanzapine of the patterns of expression of 5-HT2A and D3 receptors in rat central nervous system and blood cells. Neuropharmacology 2006; 51 (4): 923-932.
- 54. Liang CS, Ho PS, Chiang KT, Su HG 5-HT2A receptor -1438 G/A polymorphism and serotonergic antidepressant-induced sexual dysfunction in male patients with major depressive disorder: a prospective exploratory study. J Sex Med. 2012; 9 (8): 2009-2016.
- 55. Fernandes BS, Gama CS, Ceresér KM, Yatham LN, Fries GR, Colpo G, de Lučena D, Kunz M, Gomes FA, Kapczinski F. Brain-derived neurotrophic factor as a state-marker of mood episodes in bipolar disorders: a systematic review and meta-regression analysis. J Psychiatr Res 2011; 45 (8): 995-1004.
- 56. Fernandes BS, Berk M, Turek CW, Steiner J, Goncalves CA. Decreased peripheral brain-derived neurotrophic factor levels are a biomarker of disease activity in major psychiatric disorders: a comparative meta-analysis. Mol Psychiatry 2014; 19 (7): 750-751.
- 57. Lin PY. State-dependent decrease in levels of brain-derived neurotrophic factor in bipolar disorder: a meta-analytic study. Neurosci Lett 2009; 466(3): 139-143.
- 58. Kopeček M. Kognitivní efekt antidepresiv. In: Preiss M, Kučerová H a kol. Neuropsychologie v psychiatrii. Vydání 1. Praha: Grada Publishing 2006: 305-331.
- 59. Roses AD. Apolipoprotein E genoty-ping in the differential diagnosis, not prediction, of Alzheimer's disease. Ann Neurol 1995, 38: 6-14.
- 60. Schmitt K, Holsboer-Trachsler E, Ec-kert A. BDNF in sleep, insomnia, and sleep deprivation. Ann Med 2016; 48 (1-2): 42-51.
- 61. Sen S, Duman R, Sanacora G. Serum brain-derived neurotrophic factor, depression, and antidepressant medications: meta-analyses and implications. Biol Psychiatry 2008; 64 (6): 527-532.
- 62. Rybakowski JK. Response to lithium in bipolar disorder: clinical and genetic findings. ACS Chem Neurosci 2014; 5 (6): 413-421.
- 63. Björkholm C, Monteggia LM. BDNF - a key transducer of antidepressant effects. Neuropharmacology 2016; 102: 72-79.
- 64. Seifuddin F, Mahon PB, Judy J, Pirooznia M, Jancic D, Taylor J, Goes FS, Potash JB, Zandi PP. Meta-analysis of genetic associationstudies on bipolar disorder. Am J Med Genet B Neuropsychiatr Genet 2012; 159B (5): 508-518.
- 65. Harrisberger F, Smieskova R, Schmidt A, Lenz C, Walter A, Wittfeld K, Grabe HJ, Lang UE, Fusar-Poli P, Borgwardt S. BDNF Val66Met polymorphism and hippocampal volume in neuropsychiatric disorders: A systematic review and meta-analysis. Neurosci Biobehav Rev 2015; 55: 107-118.
- 66. Desai P, Nebes R, DeKosky ST, Kamboh MI. Investigation of the effect of brain-derived neurotrophic factor (BDNF) polymorphisms on the risk of late-onset Alzheimer's disease (AD) and quantitative measures of AD progression. Neurosci Lett 2005; 379 (3): 229-234.
- 67. Boiocchi C, Maggioli E, Zorzetto M, Sinforiani E, Cereda C, Ricevuti G, Cuccia M. Brain-derived neurotrophic factor gene variants and Alzheimer disease: an association study in an Alzheimer disease Italian population. Rejuvenation Res 2013; 16 (1): 57-66.
- 68. Zhang L, Fang Y, Zeng Z, Lian Y, Wei J, Zhu H, Jia Y, Zhao X, Xu Y BDNF gene polymorphisms are associated with Alzheimer's disease-related depression and antidepressant response. J Alzheimers Dis 2011; 26 (3): 523-530.
- 69. Manukyan A, Jirák R. Vztahy mezi některými genetickými polymorfismy a klinickými rysy u Alzheimerovy choroby. Česká a slovenská psychiatrie 2015; 111 (5): 236-240.
- 70. Kampinga HH, Craig EA. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nature Reviews Molecular Cell Biology 2010; 11:579-592.
- 71. Goate AM. Genetics of Alzheimer's disease; where are we now? The News, Dementia Issue 1996, 3: 1-2.
- 72. Rebeck GW, Reiter JS, Strickland DK, Hyman BT. Apolipoprotein E in sporadic Alzheimer's disease: Allelic variation and receptor interactions. Neuron 1993, 11: 570-580.
- 73. Adams C: Alzheimer's disease research: A game of connect the dots. Gerontology 1997, 43: 8-19.
- 74. Patočka J, Tichý M, Řípová D, Jirák R. Apolipoprotein E jako rizikový faktor demence. Psychiatrie 1998; 2, 108-111.
- 75. Zvěřová M, Jirák R, Fišar Z, Kitzlerová E, Hroudová J, Raboch J. Význam kortizolu a homocysteinu v diagnostice Alzheimerovy demence - výsledky výzkumného záměru. Česká a slovenská psychiatrie 2015; 5: 222-227.
- 76. Kitzlerová E, Fišar Z, Zvěřová M, Jirák R, Hroudová J, Raboch J. Význam homocysteinu a kortizolu v diagnostice depresivní poruchy - výsledky z výzkumného záměru. Česká a slovenská psychiatrie 2015; 6: 275-281.
- 77. Pláteník J, Fišar Z, Buchal R, Jirák R, Kitzlerová E, Zvěřová M, Raboch J. GSK3p, CREB, and BDNF in peripheral blood of patients with Alzheimer's disease and depression. Prog Neuropsychopharmacol Biol Psychiatry 2014; 50: 83-93.